|
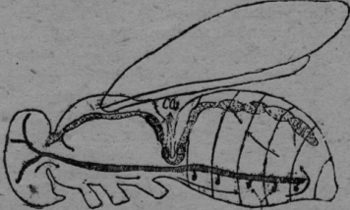
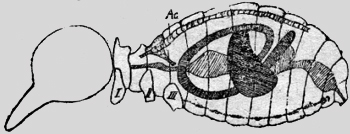
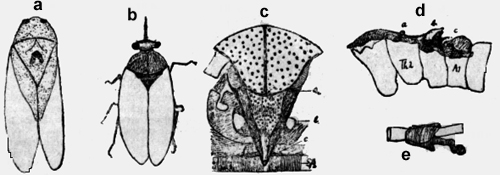
Una de las manifestaciones más características y más importantes de la vida, es su metabolismo, es decir la facultad de asimilar y desasimilar. Esto significa, que las células pueden absorber el oxígeno y expulsar el ácido carbónico. Este cambio de gases se llama "respiración interna", que es la única en los animales primitivos como los Protozoa y Coelenterata; aquí el cambio de gases se produce tras el tegumento externo. En los animales más diferenciados hay también una "respiración externa", que no es otra cosa, que un medio de transporte para integrar el oxígeno a las células y eliminar el ácido carbónico nocivo. En los vertebrados son las branquias y los pulmones, que cumplen con esta función usando la circulación sanguínea como medio de transporte. En los órganos de la respiración (pulmones y branquias) el oxígeno del aire o del agua se combina en forma inestable con la hemoglobina de los eritrocitos y puede ser retirado fácilmente de las células, mientras el ácido carbónico se difunde en el plasma sanguíneo y es eliminado por los alvéolos pulmonares o las branquias. Según la opinión comúnmente reinante hasta ahora, las tráqueas de los insectos llevan el oxígeno directamente hasta los tejidos y retiran de allí el ácido carbónico. Así escribe Berlese página 809 Vol. I, 1909: "Lo característico de los Artrópodos terrestres es la existencia de un sistema especial, por el cual el aire es llevado directamente a los tejidos y a menudo a las células mismas, así que no hay la necesidad de substancias intermedias en el transporte del oxígeno y de los otros gases, substancias como son en particular los pigmentos respiratorios". "Los insectos no necesitan un órgano de transporte para transportar de uno a otro lado el oxígeno y el ácido carbónico, como ese caso en los vertebrados con el sistema sanguíneo". (Escherich 1914, pag. 71, Forstinsekten Mitteleuropas). En mi segundo estudio mirmecológico (Acta zool. Lilloa, II, 1944) he demostrado la imposibilidad física de este proceso, de transportar dos gases en un único sistema de tubos en dos direcciones opuestas. El oxígeno del aire es aspirado por la extensión del abdomen a través de los estigmas. Después del cierre de las válvulas en el estigma, el aire es propulsado hasta la última célula por la contracción abdominal. Este es un mecanismo típico de una bomba aspirante e impelente. Pero es muy difícil imaginarse, cómo al revés el ácido carbónico puede ser aspirado en las sutilísimas terminaciones de las tráqueas; y si esto fuera exacto habría en las tráqueas una mezcla de gases, que empujada de uno y otro lado sería incapaz de cumplir con su función. Además de estas consideraciones teoréticas he llegado en mis investigaciones sobre las hormigas a los siguientes resultados positivos. Los tres estigmas anteriores dorsales poseen una estructura anatómica diferente de los estigmas ventrales del abdomen. No tienen válvulas, ni cribas y no están en comunicación con el sistema traqueal, sino con el vaso sanguíneo dorsal, la aorta, que rodean con una cámara de aire. Por fin he hecho un experimento crucial y definitivo. He tapado con goma los tres estigmas dorsales de Camponotus rufipes, Acromyrmex silvestre y lobicornis y los animales se han muerto en brevísimo tiempo (1-3 minutos) con los mismos síntomas de envenenamiento, que si se pusiera en un frasco de cianuro potásico, (temblor de las antenas y de los músculos de las piernas). Hago resaltar, que la cabeza, las piernas y sobre todo, el abdomen con sus estigmas quedaron completamente intactos, así que no puede ser la escasez de oxígeno la causa de la muerte, sino la retención forzosa del ácido carbónico. Sobre esta base he propuesto en el mencionado estudio la siguiente tesis: La introducción del oxigeno se hace en las hormigas por el sistema traqueal, el ácido carbónico se difunde en el plasma y es transportado por la sangre en el sistema vascular hasta que es expelido por las chimeneas de los estigmas dorsales. A estos estigmas dorsales los llamaré en adelante "spiracula" o espiráculos. Había observado, que si se pone una hormiga en agua, surgen burbujas de aire de estos espiráculos. Mi amigo, el Dr. Vellard, sugirió el análisis de estas burbujas con agua de bario. He puesto los animales en tubitos delgados, así que no podían darse vuelta. La cabeza y el tórax eran sumergidos en el agua de bario, quedando afuera el abdomen. El agua de bario se enturbiaba al poco tiempo. Las abejas, avispas, coleópteros, libélulas, saltamontes y moscas (Musca doméstica, Calliphora y Lucilia) daban pruebas positivas. Los animales quedaban 1/4-1 hora así en el agua de bario y quedaban después perfectamente normales, cuando eran puestos en libertad. Con las hormigas el experimento no fue tan convincente, porque estos animales eran demasiado pequeños y podían darse vuelta en el tubo, así que también se mojaba el abdomen en el fluido. Naturalmente también aquí el agua de bario se enturbiaba fuertemente. No hay duda que las burbujas de los espiráculos dorsales consisten de ácido carbónico. Me di cuenta en seguida, que este proceso fundamental de la respiración, como lo he expuesto en la tesis que sostengo, no podía ser limitado a las hormigas, sino que todos los insectos deben poseerlo. La siguiente investigación prueba, que todos los Atelecerata (Myriapoda e Insectos) poseen un aparato distinto, que permite la evacuación del ácido carbónico del vaso sanguíneo dorsal, la aorta. Estoy convencido que existe en los insectos una participación del sistema vascular sanguíneo como órgano de transporte, exactamente como en los Vertebrados, con la sola diferencia de que el oxígeno es transportado por las tráqueas. En mi ya mencionado estudio sobre la respiración de las hormigas he publicado un dibujo con un esquema respiratorio. En vez de reproducirlo aquí he hecho un análogo de la abeja (Apis mellifera), que está basado en esquemas hechos en observaciones, que no dan lugar a dudas. (figura 1). El sistema traqueal está representado por un tubo simple y anillado, que recibe el oxígeno (02) por los estigmas. Naturalmente son omitidos los sacos de aire, tubos de reducción, todas las ramificaciones y válvulas, etc. El sistema vascular está punteado. El corazón propulsa la, sangre en la aorta hasta la proximidad de la cabeza. De allí la sangre se va a la cabeza ya tórax y regresa debajo del septum ventrale en el abdomen, para ser aspirado por el corazón a través del septum dorsale. Las flechas indican el sentido de la circulación. (He aprovechado un esquema de Zander, no habiendo estudiado yo mismo este asunto).
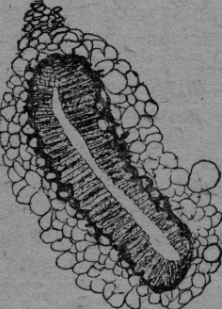
Naturalmente he observado la aorta en la profundidad de los espiráculos, que no tienen válvulas ni cribas y son solamente protegidos por su posición y cerdas. El espiráculo posterior está situado en el primer segmento abdominal en la profundidad de la cisura del cuerpo. Corresponde al estigma en el pecíolo de la hormiga. Su abertura es triangular. Dos filas de largas pestañas plumosas en la incisión del cuerpo golpean rítmicamente y llevan el ácido carbónico como un abanico a otra parte. Si se sumerge una abeja en agua, se puede observar muy bien el golpear de las cerdas y el salir de las burbujas de aire, que están formadas de ácido carbónico como prueba el agua de bario. El espiráculo mediano (Epinotum de la hormiga) está en la parte opuesta de la incisión del cuerpo. El espiráculo anterior es el más grande, como el de Metanoturn de la hormiga. Son posteriores a la inserción de las alas. Es una hendidura oblonga recurvada, que se puede ver desde el exterior del animal con un lente de gran aumento. El margen está orlado de un grueso anillo de quitina con cerdas. Pero no he podido constatar, si se mueven rítmicamente como las posteriores, porque he estudiado este espiráculo en el animal disecado y preparado. Sumergiendo la abeja en agua aquí también se ve burbujas de aire. En el esquema figura.1. tres flechas rayadas indican la salida del ácido carbónico (C02) de la sangre. El experimento con la goma no es tan claro en las abejas como en las hormigas. El tórax es muy áspero y lleno de pelos, molestan las alas y hay que cuidar que el animal no se quite la goma con las patas. La abeja se murió en diez minutos con temblor de las antenas y patas, pero a veces algunas permanecían media hora vivas. Quizá el aire de reserva en los grandes sacos traqueales juega un papel importante en esta resistencia. El experimento con agua de bario es al contrario muy claro y decisivo con las abejas. Sin duda una investigación microanalítica del bario es posible y se podría constatar cuantitativamente con este método el metabolismo del ácido carbónico. Las avispas tienen también tres espiráculos, dos en el tórax y un par cefal en el primer segmento abdominal, muy fino y alargado. La abertura espiracular es redonda, de posición laterodorsal y está protegida por cerdas. Están situados en la parte más angosta del fino pedúnculo, la aorta puede eliminar fácilmente el ácido carbónico por ambos espiráculos. Es muy interesante la comparación con la abeja. Este animal grueso sin pedúnculo acentuado posee un único espiráculo mediano, resultado de la fusión de los pares primitivos. La función respiratoria de la aorta mediana impar ha provocado este cambio morfológico. Después de describir estos hechos en las abejas y avispas, creo que todo lo que he dicho para las hormigas vale también para la generalidad de los hymenópteros. El método de investigación fue generalmente así: El animal vivo era sumergido en agua, a fin de observar con un lente de aumento de dónde surgían las burbujas de aire. He fijado los objetos en alcohol o formol o Carnoy y los he puesto en glicerina "in toto" o cortados y desmenuzados con aguja. A veces he teñido con Boraxcarmín o Haematoxylina para identificar el vaso sanguíneo dorsal. Cuando sabía el sitio y la estructura de los espiráculos, probé de obturarlos con goma y ver si los animales morían rápidamente o bien sobrevivían, poniéndolos un tiempo en agua para disolver la goma. Arriba he descrito la prueba con agua de bario, que naturalmente es imposible hacer con animales demasiado chicos (pulgas) o bien puramente metaméricos como Scolopendra y Lepisma. Las más de las veces he tomado dos representantes de cada orden de los insectos. La determinación de la especie no me fue posible en muchos casos, ni me parece importante. Muy sensible es la carencia de una biblioteca especializada, pero hay que soportar esta falta con resignación., A pesar de todo no quiero dejar de trabajar científicamente. Tengo l libro fundamental de A. Berlese "Gli Insetti" y los de Costa-Lima, Escherich, Forel y Wheeler, libros que contienen los datos generales, y ruego a los colegas excusar mis omisiones. Myriapoda En el ciempiés, una Scolopendra de 4-5cm. he encontrado el sistema respiratorio más elemental. Pero aquí también no falta el aparato de escape para el ácido carbónico en relación con la aorta. El animal tiene en cada segmento un par de pies, así que no hay una división entre tórax y abdomen. Solamente pocas partes son fuertemente quitinizadas y amarillo-café. La mayor parte del esqueleto está compuesta de quitina hyalina; solamente las articulaciones y la última articulación de las extremidades son de color café. Así es fácil distinguir a simple vista en cada segmento los estigmas amarillos, arriba de la inserción de las piernas. También en la cabeza caudal existen estigmas pares. La abertura exterior del estigma es una hendidura de 1mm de largo y está circundada como ya he dicho, de un margen quitinizado amarillo. Este "peritrema" no es liso, sino parece ser fundido de unos cuantos arcos recurvados de quitina, así que emergen en el interior picos y puntas más o menos agudas. (figura 2a y 2b).
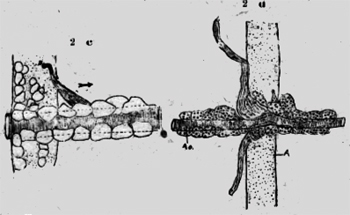
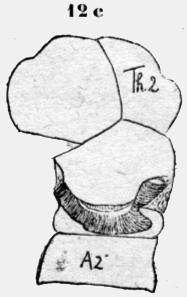
Cuando este margen de quitina está isolado, se puede observar en el interior varias filas de bastoncitos. El aspecto es similar en una retina. Estos bastoncitos tienen 30 micras largo y 5 micras ancho, están espesamente apretados y forman así un cojín plástico. No parecen de materia quitinosa, porque tienen una estructura granulosa. Creía a primera vista, que fueran células secretorias. Pero éste no es el caso. Son un estrato plástico, que empujado contra la pared opuesta, forma un cierre primitivo del estigma. A plomo de la hendidura del estigma corren los músculos estriados del cuerpo, que pueden evidentemente cerrar con su contracción la abertura externa del estigma. De la abertura del estigma se llega a una grande cavidad ovoide, que tiene una pared de chapas largas y redondas de quitina clara, pero es más espesa y fuerte que la demás cutícula del cuerpo, que muestra una escultura muy fina. En la profundidad del estigma hay caudo-lateral una larga abertura, que conduce a un saco gris, flojo y rico de pliegues. Esto se puede observar bien en una preparación de adentro. Este saco está fijado con diversos paquetes de fibras a un listón de quitina clara (figura 2a). A la -superficie del saco se ve fibras circulares y probablemente son músculos. Pero no he pedido constatar una estriación transversal y no se si existen músculos lisos en este animal.. En un preparado de un estigma isolado se ve muy distintamente, que músculos estriados provenientes de adentro se insertan en el saco (figura 2a). Aparentemente estos músculos dilatan el saco respiratorio por la inspiración. Probablemente el aire en el saco es expulsado por las fibras circulares arriba mencionadas, pero también los músculos del cuerpo, que corren a lado del saco, pueden participar en este asunto. Me parece más probable, que las fibras mencionadas circulares son contráctiles, porque he visto en estos animales primitivos tal vez músculos indudables sin estriación perceptible. En la profundidad del estigma opuesto a la abertura exterior existe una ranura hueca, de la cual parten las tráqueas de gran calibre en todas las direcciones. Algunas tráqueas poseen especimientos globosos, estrechamientos en forma de embudo, pero mucho menos que en las hormigas. Un tronco longitudinal no existe en el sistema traqueal de este animal. Tampoco se encuentra una válvula regular, como en los demás insectos. He hecho diversas preparaciones con dos agujas bajo de una lente de aumento bastante fuerte y no creo, que ninguna válvula se habría escapado a mi observación si existiera. Así no queda más que dicho cojín de bastoncitos como aparato para cerrar el estigma. Los músculos antes mencionados empujan el aire encerrado en el estigma y el saco por el paquete de tráqueas en el cuerpo. Un sistema traqueal más simple no se puede imaginar. Llama la atención el tamaño de la hendidura y del estigma mismo, como el gran calibre de las tráqueas. Aparentemente este aparato muy primitivo funciona mediocremente y la cualidad es reemplazada por la cantidad. Cada segmento posee a cada lado un estigma y con las tráqueas colosales el arribo de oxígeno será. bastante. Con el perfeccionamiento del Aparato respiratorio se reducen el número de los estigmas y el tamaño de las tráqueas. Posiblemente el saco respiratorio es el- único propulsor del aire y no la contracción de los segmentos del cuerpo. Porque no hay segmentos abdominales. Cada uno tiene también sus piernas y no existe una; separación del tórax y abdomen. Nunca he observado una contracción entre los segmentos, como se ve en los insectos, en el abdomen. Es difícil imaginarse cómo el ácido carbónico pueda ser evacuado "al revés" con este mecanismo respiratorio de escolopendra. Esto naturalmente no es necesario, porque existen aparatos especiales para permitir el escape de este gas de la aorta.
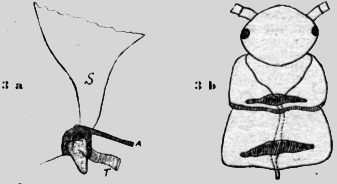
Estos aparatos son dibujados de adentro en las figuras 2c y 2d. El vaso sanguíneo dorsal está orlado por grandes células, que evidentemente corresponden a células hepáticas con sus vacuolas y trozos de chromatina. Se encuentran también en otros lugares del cuerpo Poco antes del margen caudal de un segmento, parte de la aorta un saco de quitina hyalina con base larga, que se estrecha mucho en su transcurso caudo-lateral, para terminar en un puño redondo, fijado en la cutícula exterior del cuerpo. En esta chimenea de escape no he visto células, pero sí una rayadura longitudinal muy fina. En. la figura 2c he dibujado también la burbuja de aire, que indicaba distintamente el fin del tubo de escape. He preparado y aislado este tubo de ventilación (figura 2d). En: un lado se ha roto y en el otro está desgajado, pero, muestra muy bien el puño redondo terminal y la delicadeza de su estructura. En esta figura 2d se ve también, cómo el vaso sanguíneo está abrazado por las bases largas del tubo de escape. Poco más craneal de esta parte ensanchada de la aorta, hay un .rodete en el vaso sanguíneo, que ciertamente tiene una :función regulatoria; sea que cierre el vaso y ataje la sangre para el escape del ácido carbónico, sea que sirva como corazón segmental para propulsar la sangre. Ningún experimento con goma o agua de bario es posible con este animal, que posee todo en cada segmento. Apterygota Thysanura Lepisma. Carl W. Neumann escribe de los insectos primordiales en el Brehm 1929 en la página 519: "Las aberturas respiratorias desembocan en tubos, que se reúnen solamente en pocas especies en un sistema (traqueal) y además se encuentra las más veces ventral, sacos respiratorios chicos en los anillos abdominales, que las más veces quedan retirados, pero pueden también surgir. Si la vecindad es muy húmeda estos sacos se llenan con aire y emergen mas o menos". Puedo confirmar estos hechos y -tengo una preparación con todos los sacos afuera llenos de aire. Mi objeto de investigación fue un "pececito de plata" un Lepisma saccharina de 10-14mm. de largo. El objeto es bastante difícil de estudiar. Especialmente estorban las escamas brillantes con sus rayas a distancia de tres micras unas de otras. Existen ya descripciones del aparato respiratorio de este animal y me limito a unas cuantas observaciones, que me parecen de importancia para mi asunto. El estigma está fijado al anillo segmental posterior. No es redondo, sino un poliedro, oscuro y cubierto de escamas. El estigma tiene tres aberturas: una hendidura chica y estrecha, que comunica con el exterior, una segunda caudal va al saco respiratorio y una tercera medial por las tráqueas. Delante de esta abertura está una membrana, que aparentemente puede cerrar la abertura como una. válvula. (figura 3a). El saco respiratorio se compone de una membrana delgada de quitina con puntos oscuros. Un múscúlo proveniente del interior se inserta al saco y puede rétirarlo. El relleno con aire debe producirse pasivamente con respecto al saco. También no existen fibras circulares, como en la Scolopendra. Así parece que sea un saco de reserva de aire y no un órgano de ventilación. Lepisma está más diferenciada que-la Scolopendra,posee solamente tres pares de extremidades funcionantes (las otras son rudimentales). Así hay la sepáración de un tórax de un abdomen,y este abdomen puede asumir la función de la inspiración. Los sacos respjratorios. atávicos asumen una función secundaría y se pierden después, en 1ª filogenia en los demás insectos.
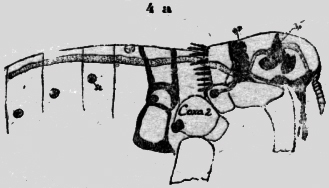
La cosa principal para mí son los órganos dorsales para la ventilación del ácido carbónico. Son muy similares a los de Scolopendra. En la figura 3b he esbozado los dos segmentos detrás de la cabeza. Se puede observar en la parte caudal del segmento en ambos lados de la aorta un saco hyalino, que desemboca con un puño estrecho en un lugar libre de escamas. El dibujo está `combinado de diversos preparados, porque en uno he podido observar los tubos de escape, y en otro la aorta serpenteante, cuando las escamas eran eliminadas. En todo caso este insecto primordial posee un, aparato metamérico. Dorsal para evacuar el ácido carbónico transportado por la sangre en la aorta. Aphaniptera La pulga del perro, Ctenocephalus canis (Curt.), posee un aparato respiratorio extremamente curioso y singular. La posición de los estigmas ya no es común. Los más caudales están situados abajo en la región ventral, mientras los siguientes cefal siempre más dorsal, hasta que el primer segmento abdominal posee un único estigma impar en la línea mediana dorsal (figura 4a).Los estigmas son cúpulas chatas redondas con una abertura oval al lado. En la parte opuesta desemboca la traquea con una larga abertura redonda. En el interior del estigma están: tres pares de bastoncitos de fuerte refracción, que convergen hasta el centro. Arriba de este armazón yace un grumo gris, que parece como una esponja. Esta masa llevada por los bastoncitos está situada en medio de la abertura exterior y la tráquea. Con la dilatación del abdomen el aire puede ser aspirado libremente, pero con la contracción del abdomen esta masa será echada contra la abertura externa y la tapará. Esta es una verdadera válvula automática sin músculos, que regula la bomba aspirante e impelente. Está excluida con este aparato la posibilidad de evacuar el ácido carbónico "al revés" (figura 4b).
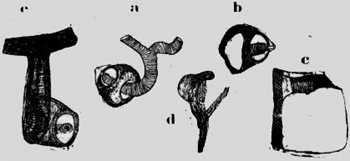
De los estigmas abdominales van paquetes de tráqueas bastante gruesas a los órganos. Un tronco longitudinal no existe, Pero las tráqueas comunican entre si con anastomosas intersegmentales oblicuas. Después de salir del estigma la traquea ancha tiene un tubo muy delgado sin hilo espiral; pero con dos bolsas laterales. Este tubo en forma de embudo intercalado es una medida de seguridad para el estigma, cuando con la contracción abdominal el aire es propulsado en el interior del cuerpo por presión. Luego este tubo expansible se. pone hueco como un globo de goma y protege el estigma de una presión excesiva.(figura 4b). En el sistema traqueal están interpolados numerosos sacos respiratorios. Por ejemplo en la tibia pasa una tráquea delgada, con rígido hilo espira1 a un saco extenso, del cual surge otra tráquea delgada, pasa la articulación y desemboca en un saco en el tarso. He podido observar en el microscopio con leve presión del cubre objeto, cómo el aire del saco tibial pasaba al saco tarsal. Sin duda estos sacos son dilatables y también contráctiles por un estrato fino muscular. La pulga posee, además de los estigmas abdominales, un estigma en cada segmento toracal, situado caudo-dorsal en la coxa. Se pregunta con asombro, cómo es posible técnicamente una ventilación de un estigma puesto en la coxa rígida y fija, que naturalmente no puede dilatarse. Una, investigación más exacta ha dado la explicación, que a primera vista parece fantástica e, increíble si no habría analizado cada uno de los componentes. Se trata de una bomba aspirante e impelente regular con un émbolo en un cañón sólido, que es empujado por músculos de una y .otra parte. En la figura 4c están dibujadas exactamente cada una de las: partes aisladas que componen la bomba; el conjunto es forzosamente esquematizado.
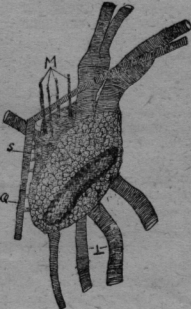
Los estigmas de la coxa son totalmente diferentes de los abdominales. Tienen la forma de un cáliz alto, de calor, café oscuro con abertura espaciosa cubierta de un techo de bóveda transparente como el vidrio. Aquí está una abertura lateral. De adentro entra una tráquea espaciosa y termina con un botón redondo, sobre el cual está sobrepuesto un tubo hyalino. Aparentemente este tubo intermediario claro corno vidrio establece la comunicación con la abertura. externa y es un mecanismo para cerrar, si la presión aumenta, quizá retorciéndose. En todo caso el aire puede entrar sin obstáculo en la inspiración y es retenido en la "compresión" y empujado hasta los órganos interiores del cuerpo. En la cápsula del estigma: existe una pared intermedia transversal, por la cual pasa la tráquea, que se fija con aquella. En la base del estigma se encuentran dos agujeros en diferente enfoque microscópico. Por el más alto y lateral sale la tráquea (figura 4Ca), mientras el otro mayor y oval (figura 4Cb), se pone encima de un tubo de quitina obscuro con pared: muy gruesa,(figura 4Cc). En este cañón se encuentra un émbolo de forma extraña,(figura 4Cd). Es un bastón de quitina fuerte recurvado al cual se sobrepone una parte redonda de quitina más clara, como un sombrero. Quizá es una masa plástica, que rellena bien la cavidad del cañón, pero permite el movimiento del pistón. Al arco macizo se insertan dos músculos estriados, que pueden tirar el émbolo en una dirección y otra. En la figura 4Ce. todo. este aparato está reunido esquemáticamente. Cuando el músculo tira el émbolo hasta el margen sólido de la coxa, el aire es aspirado por el estigma en el cañón, si el músculo antagonista retrae el embolo, la válvula del estigma dirige el aire al interior del cuerpo por las tráqueas. Así esta bomba: es suficiente por sí misma y totalmente independiente de la contracción abdominal La distribución del aire es, controlada por los sacos respiratorios intercalados en el sistema traqueal. Los de las piernas están ya mencionados. En la figura 4a se ve dos grandes sacos en la cabeza, la cual es provista con oxígeno del estigma de la coxa I. Una gruesa tráquea va de este estigma al saco respiratorio situado al margen caudal de la cabeza. Otra tráquea lo conecta con un segundo saco debajo del ojo. Los sacos respiratorios son como ya he dicho, contráctiles (en las abejas he observado fuertes músculos longitudinales). De este modo el arribo de oxígeno por los órganos de la cabeza es claro y comprensible. Escherich cita la pulga como animal que posee excepcionalmente un estigma en la cabeza. Pero. este órgano no funciona por 1a inspiración, sino es un espiráculo, que sirve solamente para la evacuación del ácido carbónico. No tiene ninguna conexión con el sistema traqueal. Este espiráculo está situado dorso-caudal del ojo, en un triángulo formado por listones de quitina. Al lado del espiráculo se extiende una membrana granulosa, que cubre un vasto y profundo,. espacio hueco. Este se abre más caudo-ventral por afuera, pero está cubierto por dos filas de cerdas, que se cruzan en ángulo recto. Muy claro se puede distinguir el vaso sanguíneo directamente bajo del espiráculo (figura 4a). El ácido carbónico puede salir de la aorta una vasta cúpula y evacuar al lado por la estacada de los pelos. Una segunda posibilidad de escape existe para el ácido carbónico, más ventral en el límite entre la cabeza y el tórax. Aun aquí existe un triángulo de quitina. En la profundidad corre el vaso sanguíneo, que-para este; fin tiene una curvatura aguda. Cerdas gruesas forma en la superficie una cubierta de protección. La cabeza de la pulga es muy delgada y así es posible, que el ácido carbónico salga de ambos lados del Sanguíneo en la profundidad de las hoyas expiratorias. En el margen caudo ventral de las- cabeza, el tórax tiene un planos, pendiente, así que se forma una quilla ventral, que permite también la evacuación del gas por ambos lados. De aquí se ve, como la función respiratoria modifica y modela la estructura morfológica. En comparación con los Hymenóptera, Pulex posee un sistema respiratorio más primitivo. Sus estigmas son "metamer" en cada segmento como en Scolopendra. Falta un tronco longitudinal. Su estigma tiene una válvula automática sin músculos. Como a menudo en grupos relictos sobrevivientes, que no, se acomodan al sistema común, se encuentran en la pulga órganos singulares como los estigmas en la coxa. Recuerdo los Gymnophiones Stegocephales sobrevivientes, con sus tentáculos, escamas, calcáreas en la piel y los corazones segmentales linfáticos. En los siguientes órdenes de los insectos he presumido el sistema traqueal como conocido y no lo mencionaré sino excepciónalmente. Describiré principalmente el aparato expiratorio en conexión con el vaso sanguineo dorsal. Principio con el grupo de los isoptera. Isoptera He investigado una Termes del Temporál de Cochabamba con soldados Nasuti de 3 mm, ninfas aladas de 7 mm, y trabajadores de 2,5 mm de largo. Este Eutermes sp. posee el escape para el ácido carbónico en el limite entre el abdomen y el tórax. En la figura 5 he numerado coxas. Dorsal en el tercer segmento se ve una espaciosa cámara de aire abrazar la aorta. Este saco (punteado) se estrecha en un tubo delgado, que desemboca lateralmente en el primer segmento abdominal por afuera.
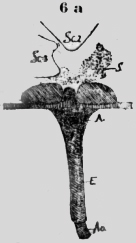
El sistema intestinal es rayado, el nervioso es punteado. En los trabajadores de la misma especie hay hechos análogos y tambier en un soldado de l2 mm de San Ignacio (Chimóré) con largas mandíbulas recurvadas. Orthoptera Acrididaé. He investigado una serie de langostas y he observado en todas hechos análogos, sea en un Stenobrotus o una Eremobia con alas rojo negrás o en la langosta gris con alas amarillas, tan común aquí con su sonido estridente. En la línea mediana dorsal se realza craneal una cumbre, la cual desciende en un plano pendiente al límite con el tórax, ; Aquí hay: una abertura triangular (figura 6 a).
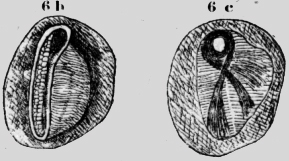
En la profundidad de la cumbre corre la aorta, que parece terminar en la región toracal. En el espacio del "scutellurn" impar y los escudos laterales del tercer segmento se ve la sangre amarilla difusa. Todo el espacio hueco debajo de la cumbre es hueco y sirve de chimenea para el ácido carbónico, que se evacua adelante por la abertura. He tapado ésta abertura con goma, pero el animal sobrevivió. Esto lo comprendí cuando vi dos espiráculos pares detrás de cada ala, que corresponden a los espiráculos del Epi-y Metanotum, en las hormigas. Grillus tiene un aparato análogo, pero la cumbre es más baja. Quiero describir su estigma, porque tiene un, modo de cerrarse singular. Los estigmas están situados en medio de cada metámero en una faja no segmentada. Están hundidos en una cavidad redonda pigmentada. La hendidura tiene 200 micras de largo y es perpendicular al eje del cuerpo. El estigma es claro y tiene la forma de una bolsa chata de tabaco con cierre a cremallera. La hendidura está circundada de un margen de quitina con cerdas cortas y adentro un cojín con chapas. Débiles músculos transversales pueden abrir la hendidura del estigma en la fase de inspiración (figura 6b) Si se observa el estigma al revés desde adentro, se ve surgir en un lado un tubo bastante angosto (70 micras), pero muy sólido. Alrededor de este tubo corre un lazo de músculos fuertes, que se insertan al lado del estigma (figura 6c). Si estos músculos se contraen, tiran el tubo mencionado hasta el centro del estigma y con esto cierran la abertura exterior, como uno se puede convencer fácilmente con el modelo de la bolsa de tabaco. Al tubo angosto se insertan tráqueas gruesas de un calibre de 250 micras. Phasmidae He investigado dos especies (Diapheromena?). Una de Cochabamba (la; hembra de 7cm. y el macho de 4cm., de largo). El color varia de amarillo paja al verde y negro. La otra especie proviene- de Santa Cruz el "mata-caballos" de 8-10cm. de largo y de un verde oscuro metálico.
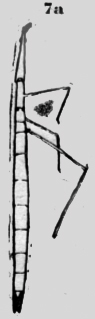
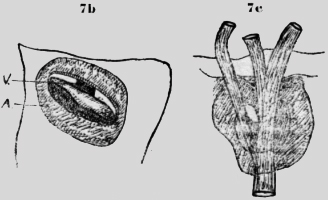
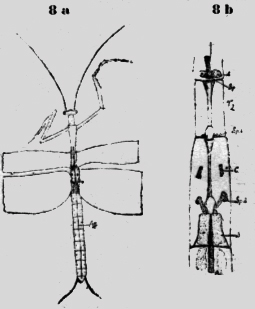
Ambas especies tienen en el primer segmento toracal, vecino a la cabeza, aberturas pares, cerca de la línea mediana. Estos espiráculos dorsales están situados en una cúpula un tanto elevada, que permite la evacuación del ácido carbónico de la aorta mediana. También existen espiráculos en los dos siguientes segmentos toracales, como se ve en la figura 7a. Debajo de la abertura externa, de estos espiráculos existen laminas quitinosas transversales con hendiduras. Así el vaso sanguíneo, que corre allí en la profundidad es protegido y de otra parte las hendiduras permiten el escape del ácido carbónico. La figura 7b muestra el espiráculo, visto de afuera y la figura 7c de adentro. Oethecaria De los Oethecaria he investigado una Mantis de 6cm. de largo. Los estigmas segmentales están situados lateral y dorsal al margen del cuerpo. El primero es muy grande; mucho más que los demás. Probablemente ha de suplir toda la parte anterior. Existen elevaciones redondas con una hendidura transversal en el primer segmento toracal y en el segundo, donde queda un espacio libre entre las alas. Estas cúpulas están situadas en vecindad inmediata de -la cumbre mediana dorsal, así que también aquí la aorta puede evacuar por ambos lados el ácido carbónico. Otra especie de mantis de cinco cm de largo y nueve cm de ancho con las alas extendidas me parece similar a la Ameles de Europa reproducida en la Encyclopedde (Orthopteres)de C. Haulbert 1926. Las alas posteriores tienen una cinta roja craneal. Poniéndola en agua en un tubo, salieron, grandes burbujas de aire continuamente de debajo de las alas posteriores del espiráculo 3 (figura 8b).
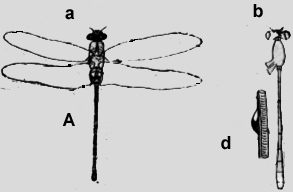
Este animal, fijado en. alcohol he investigado más exactamente que los otros ejemplares secos. Esta Mantida posee tres espiráculos que pertenecen a los tres segmentos toracales. Berlese afirma que todos los embriones poseen un estigma en cada segmento toracal. Por esto el sitio de los espiráculos en Mantis es el primordial y en los demás insectos han emigrado detrás al primer segmento abdominal: Este proceso es fácil de comprender si se considera los sacos respiratorios accesorios en la figura 8b, que están en comunicación con, la cámara de aire S y que están situados en el primer segmento abdominal. La aorta relucía "in vivo" en la línea mediana dorsal. También se ve en medio de cada segmento los estigmas (figura 8a). En el primer segmento abdominal existen al lado de la aorta dos sacos respiratorios, ya mencionados, que se unen craneal en un gran saco (S), que evidentemente abraza la Aorta, que desaparece en un surco longitudinal mediano. De la cámara de aire S parten dos tubos encorvados con aberturas laterales (Sp 3). En el medio de este tercer segmento toracal hay dos manchas negras, que a primera vista tomé por los espiráculos, pero no es así. No tienen aberturas. Son aglomeraciones de pigmento, un "camouflage" (figura 8b; C), que protege las verdaderas aberturas del espiráculo. Lo mismo vale para las dos cúpulas en el primer segmento toracal. Los puntos negros no son aberturas, sino pigmento y el espiráculo impar está situado en medio de la profundidad. (figura 8b; C y Sp. 1). En la línea mediana del segundo y tercer segmento existe un espigón hueco, que termina caudal en una elevación oval. Ciertamente es un refuerzo para el cuerpo estirado, pero siendo hueco .la aorta en la profundidad puede evacuar el ácido carbónico, que sale después de los espiráculos. Amphibiotica He investigado dos especies de Libélulas, una grande del tipo Aeschna y una Agrion de 3cm de largo. La Aeschna (7-8cm. de largo) posee en medio del primer segmento abdominal una elevación como un techo, fue forma un túnel, que se abre detrás. Al lado de esta abertura corren dos cintas negra transversales, que ocultan de este modo el agujero también negro. Al extremo caudal de este segmento hay otras cintas negras como una repetición del dibujo superior y como perfeccionamiento del "camouflage". Si se saca el techo del túnel, se ve una vasta cavidad hueca, en la profundidad de la cual corre la aorta (figura 9a). Más fácil de investigar es la delicada. Agrión (figura 9b), que tiene un cuerpo de color amarillo-verde abajo, y negro arriba. Solamente en el primer segmento existe una cinta más azul (figura. 9c), que se eleva dorsal un tanto en ángulo obtuso y se baja más craneal paulatinamente. Con esto llega en el color negro, así que también en éste caso no se puede distinguir fácilmente la abertura negra y no la he sospechado, el porqué se encontraba allí una burbuja de aire. Después de haber diseccionado el animal, he visto arriba del vaso sanguíneo, la cúpula y en el lugar donde termina el color verde azul, la abertura del espiráculo (figura 9d). Además existen dos espiráculos medial de la inserción de las alas, que están dibujadas en la (figura 9d). Vistas al microscopio, agrandadas, se observa cerdas transversales, que protegen la entrada (figura 9e).
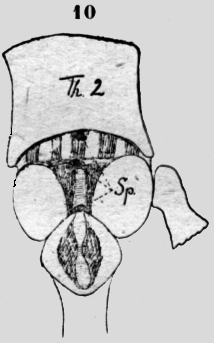
Neuroptera El Myrmeleon sp. posee tres espiráculos en la línea mediana dorsal, craneal del primer segmento abdominal (figura 10). El primer segmento abdominal es un rombo con ángulos redondos. En el centro una hendidura oval va en la profundidad, donde corre la aorta. Esta hendidura encordelada en medio, está cubierta de una membrana estriada transversalmente, pero la cavidad se abre más craneal en un .plano pendiente del segmento con una abertura oval para afuera. En el último segmento toracal una parte mediana clara está limitada por dos laterales oscuras. También en este segmento el espiráculo está situado en la profundidad craneal, sostenido por dos listones transversales (figura 10).
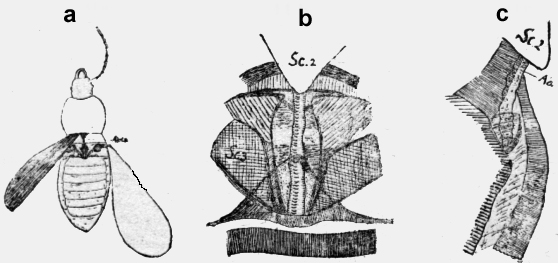
El segundo segmento toracal posee su espiráculo en un plano muy pendiente caudal. Aquí hay también una estriadura de cintas claras y negras y el espiráculo está oculto en la cinta negra central. Una Chrysopida parece tener espiráculos pares, pero no poseo más que material montado a seco y por esto no quiero afirmar nada. Coleoptera Un Carabus de 16mm. de largo, puesto en agua emite. burbujas de aire del scutellum Poniendo goma en este lugar, el animal no se muere. Probablemente los coleópteros poseen como todos los demás insectos dos espiráculos toracales, pero no he podido identificarlos en el carapax tan denso. Todos los Coleópteros poseen una fosa profunda debajo del scutellum (figuras 11a y b). Es una cavidad profunda rómbica, que está: formada por fuertes palos de quitina con los escudos del tercer segmento (Sc3). Éstos sostienen un alto andamio por las alas. En la profundidad de este hoyo está una lámina triangular de quitina (figura 11b punteada), sobre la cual yace la aorta. Si se arranca los "Elitros" aparece en la profundidad el vaso sanguíneo libre a la luz del día. La aorta prosigue abajo del secutellum (figuras 11b Sc2). Después de arrancar los Elitros de un Carabus amarillo-verde metálico, he puesto una gota de goma en esta cavidad y con una aguja, encorvada empujé la goma adelante y abajo del scutellum. El animal no fue herido; según creo, pero se murió a los tres minutos con espasmos en las piernas, esto es con signos de envenenamiento producido por el ácido carbónico retenido. No obstante este resultado positivo, no quiero afirmar, como ya he dicho, que faltan los dos espiráculos toracales en los Coleópteros. En la figura 11c se ve un corte longitudinal de una Coccinella de 9mm. de largo. La aorta libre aparece y se baja detrás del scutellum. El hoyo profundo caudal del Scutellum existe en todos los Coleópteros. Remito a la figura de :Ganglbauer de una, Cicendela (reproducida de Escherich pág. 27). No se que función los autores le han atribuido, pero es claro, que para mi es un escape para el ácido carbónico.
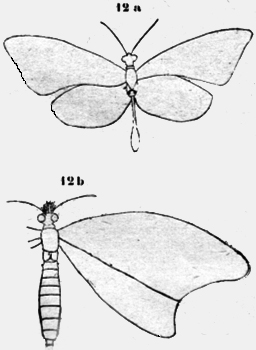
Lepidoptera Las mariposas poseen 3 espiráculos pares. Una Diurna; que me fue determinada como Athesis. confusa; tiene en el primer segmento abdominal una espesura globosa con dos hendiduras encorvadas (figura l2a). Otra Diurna amarilla, una Argentida, posee en este primer segmento dos excrecencias cónicas (figura 12b). Las aberturas laterales han sido controladas en el microscopio. En este caso no fue posible identificar con seguridad el vaso sanguíneo, pero debe correr en el medio de la cúpula hueca y el ácido carbonico puede salir de ambos lados.
Si se ahoga una-mariposa (mejor después de cortar las alas), se ve salir en este lugar burbujas de aire. Muy distantemente he podido observar la evacuación de aire en una mariposa chica como una Lycaena con alas grises y cuerpo amarillo. Bajo leve presión salió una burbuja de aire situada al centro del primer segmento abdominal de una abertura lateral detrás del último segmento toracal. El esbozo 12c ilustra esta relación. Otros dos espiráculos pares existen en el segundo y tercer segmento toracal, detrás de la inserción de las alas. Las aberturas son protegidas por largos pelos. Diptera Si se pone una gota de goma adelante de las alas de una Mosca doméstica, después de 1-2 minutos de patalear vehementemente, se queda sin reacción como muerta. Pero si se pone en este momento la mosca en agua, la goma se disuelve y largas burbujas de aire surgen de este lugar mencionado. También el animal patalea vehementemente de nuevo. El experimento con agua de bario demuestra que las burbujas de aire son el ácido carbónico.
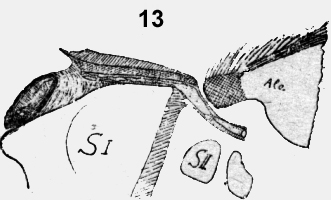
En una mosca diseccionada y aplastada entre dos porta-objetos se puede reconocer al final del primer y segundo segmento toracal un espiráculo, protegido por pelos y largas cerdas: Además se ve la aorta que corre, en un semicanal y se baja después en un túnel espacioso abajo del espiráculo (figura 13). Después de conocer estos hechos no es difícil de identificar cráneo ventral del ala una hendidura como espiráculo. La mosca no es objeto favorable y prefiero proseguir la descripción con una Nematocera, un mosquito de 4mm de largo.
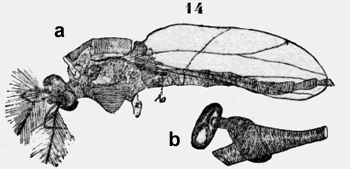
En la figura 14 he dibujado un macho de esta Nematocera y los números en la coxa indican los segmentos toracales. El espirácmlo anterior en mayor engrandecimiento demuestra el envolucro de la aorta (figura 14b). Una camera llena de aire circunda el vaso sanguíneo aquí dilatado y prosigue en un tubo con hilo espiral análogo como en una tráquea. Este tubo termina en la profundidad del espirácido. Allí existe una lámina de quitina en forma de Y como sostén y protección al mismo tiempo contra cuerpos ajenos. El margen superficial tiene una aorta de pelos. Este espiráculo pertenece al segundo segmento corresponde al Metanotum de la hormiga, y solamente la abertura está al extremo del primer segmento. El espiráculo del tercer segmento se ve en la figura 14a. La abertura del espiráculo está detrás de la inserción del ala. Finalmente hay un espiráculo impar en el primer segmento abdominal. La aorta (figura 14a Ao) corre debajo de este espiráculo, que me parece estar cubierto por una fina membrana con criba. Pero no quiero afirmar, que no exista un agujero anterior. En todo caso los Dípteros poseen un buen aparato para la evacuación del ácido carbónico . Rynchota Heteróptera. Una vinchuca, Gonorhinus infestan, tiene un profundo hoya, como Carabus. En su profundidad corre la aorta (figura 15 punteada) en el triángulo libre que es sostenido caudal por un escudo y dos vigas divergentes, que sirven de sostén para las alas. El ácido carbónico puede salir fácilmente del vaso sanguíneo pero existe ciertamente otros espiráculos toracales, por que sobrevivió el animal, cuando tapé el hoyo con goma.
Homoptera. He investigado tres especies de cicadea de Cochabamba. La más grande (3cm. De largo con alas) posee al dorso una elevación con túnel similar a la de la Aeschna, pero la abertura está craneal (figura 16a.). He investigado más en detalle una Membracida Aethalión sp. de 1cm. de largo, que posee un sólido escudo triangular anaranjado entre las alas (figura 16b). Este escudo tiene agujeros para los pelos del tacto (figura 16c) La Membracida posee los tres típicos espiráculos para el escape del ácido carbónico, que están marcado con a, b, c, en la figura 16c. En la vista de arriba no se puede sospechar el espiráculo del primer segmento abdominal en la profunda cisura entre el abdomen y tórax. Pero en un corte paramediano se ve, cómo el abdomen se baja en un plano pendiente y se puede constatar la abertura del espiráculo (figura 16d.) el vaso sanguíneo dorsal está envuelto por un espacioso saco de aire, del cuál sale un tubo de escape hasta el botón globoso terminal del espiráculo (figura 6e). No he visto ninguna válvula u órgano de protección, que no parece necesario, si se tiene en cuenta la situación protegida en que está colocado este espiráculo. El espiráculo del tercer segmento toracal está situado en la profundidad de una espaciosa cavidad, que en su parte medial tiene por techo una fina membrana, que lateral comunica libremente con afuera (figura 16cb).Por la superficie de esta cavidad, pasa oblicuamente un listón dentado muy oscuro (fuertemente quitinizado) a la extremidad caudal del Scutellum. Este listón sirve evidentemente de sostén y protección. En la figura 16c. este espiráculo b. está dibujado solamente a la izquierda, mientras en esta vista no se puede distinguirlo a la derecha.
En el corte paramediano 16d. se ve cómo un vasto tubo con hilo espiral termina al fondo de la cavidad, protegido por el techo medial. El espiráculo anterior (a en la figura 16c.) está situado al lado del Scutellum, al confín del segundo y tercer segmento toracal. Se lo ve también en la figura 16b. Termina con un botón globoso de un largo tubo de escape, que proviene de un saco en torno al vaso sanguíneo. Aquí también no he visto ninguna válvula, como en los otros espiráculos. Pero no quiero ocultar que en los estigmas no he encontrado también válvulas regulares. La hendidura oval está circundada de un margen oscuro con una orla de cerdas. Aparentemente los estigmas están cerrados por un lazo de músculos exteriores. Aphididae De estos toscos y perezosos animales he investigado un pulgón alado de 2mm. de largo, que vive sobre las flores de un filocactus. Los animales tienen un color verde, que se pierde en alcohol. Las hembras aladas eran café oscuro. Poseen tres espiráculos (a,b,c,). En el primer segmento abdominal al confín con el tórax existe un triángulo, cubierto con pelos delgados, así que parece como estriado (figura 17a). Además hay otros dos espiráculos pares, uno antes de la inserción del ala posterior, y otro detrás del ala anterior (figura 17a.). Al lado del Tergum oscuro y arqueado existen tubos delgados de escape, que tienen la forma de un ventilador encima de la cubierta de un buque
Esto se distingue claramente en la vista lateral (figura 17b), donde un tal "ventilador" está dibujado al lado en tamaño mayor. Consideraciones generales Después de haber constatado en todos los insectos sin excepción un aparato expiratorio para el ácido carbónico, creo haber cumplido mi propósito de probar mi Tesis, que la sangre es un órgano de transporte esencial para la respiración en los insectos. Pero hay que considerar otra cuestión en la respiración de los insectos, que no es de importancia para los pulmonados. Es la pregunta: ¿Qué se hace el Azoe introducido con el aire en el cuerpo del insecto? En los vertebrados el ázoe inspirado en los pulmones, se va en la expiración, inalterado, sin contacto con los tejidos. Pero en los insectos el aire es empujado hasta las células por el sistema traqueal. El oxígeno es arrancado y consumido por las células y el ázoe restante, llegado con el aire, ha de ser eliminado del cuerpo de un modo u otro. La afirmación de la condesa Linden, que las orugas sintetizan el nitrógeno del aire en albúmina, no ha sido confirmado y aceptado por los fisiólogos. Tampoco hay indicios que el ázoe sale con el ácido carbónico. Mucho más probables me parecen las siguientes vías de evacuación: A menudo se observa, que burbujas de aire salen de la boca y del ano, cuando se pone un insecto en un fluido. También se ven burbujas de aire en el intestino en preparaciones frescas en glicerina. Así me parece imaginable, que el ázoe superfluo sea eliminado por el intestino. Otra posibilidad se basa sobre la observación, de que un insecto, que se asienta después de un vuelo, mueve vehementemente su abdomen, como un acordeón, pero por breve tiempo. El animal en descanso necesita poco oxígeno en comparación con el gasto durante el vuelo. Estas contracciones del abdomen producen sin duda una ventilación de los grandes sacos respiratorios, que todos los insectos voladores poseen. Creo que con esta ventilación sale el aire rico en ázoe y pobre en oxígeno de los sacos respiratorios y que es reemplazado por aire normal. Este proceso se puede conseguir por una verdadera expiración. En estos insectos alados los estigmas están regulados por válvulas equipadas con músculos estriados, así que el aire puede ser expulsado "al revés" afuera. En Lepisma o la pulga con su válvula automática esta vía de evacuación me parece imposible y la eliminación ha de efectuarse de otro modo. Si estas consideraciones son justas, estaría clara también la función de los sacos respiratorios hasta ahora oscura y discutida. Los sacos serían receptáculos de aire para entregar el oxígeno durante el vuelo. Después en el descanso el insecto renueva el aire con el ázoe acumulado en los sacos respiratorios con aire normal por medio de rápidos movimientos abdominales. Naturalmente este proceso puede ser aprovechado comúnmente por todos los insectos sin volar, si poseen válvulas reguladas por músculos, así que el aire puede ser dirigido en ambas direcciones a voluntad. Así se comprende también la aparición de válvulas con músculos propios en los estigmas de los insectos más diferenciados, mientras los primordiales poseen válvulas automáticas. Son siempre las precisiones fisiológicas, que forman y modifican la estructura morfológica. Para la protección del animal Es claro que un agujero bastante grande, como un espiráculo, es un peligro para un animal, protegido por una firme coraza de quitina. Observando la variedad de dibujos y colores en los insectos, un hombre no poeta se extraña, que la naturaleza usualmente tan parca y realista, haya prodigado tanta belleza a primera vista inútil.. Pero un estudio atento muestra la falsedad de esta opinión. Estos colores y dibujos tienen el oficio de "camouflar" los puntos débiles de la coraza; así ocultan las Libélulas, Zánganas, Myrmeleon, Mariposas, Langostas y Cicadas sus espiráculos. La abertura negra no aparece por medio del dibujo o del color.demostrativa es la figura 9 de Aeschna y Agrión. Otro modo de protección para el animal es el lugar, en que están situados los espiráculos. Así por ejemplo las Abejas y Cigarras tienen estos puntos débiles de su coraza en la cisura profunda e inaccesible entre abdomen y tórax. Finalmente hay órganos propios de protección, como el Scutellum en los Coleópteros, listones dentados superficiales (Aethalion) o lámina profunda (Phasma y Diptera), que protegen la entrada al espiráculo. Otra vez el escape está al lado como en las pulgas y pulgones, o bien un túnel conduce afuera (Libélula, Cicada). Las cerdas y pelos, que orlan la abertura no protegen al animal, sino al aparato respiratorio de cuerpos ajenos como mugre. En pocas palabras se puede afirmar, que no existe un espiráculo que no esté oculto de un modo u otro. Esta es quizá la razón, por qué hasta ahora estos aparatos respiratorios no han sido reconocidos en su valor real. La Filogenia del sistema respiratorio 1) La respiración primordial se produce sin órganos especiales, como en Protozoa y Coelenterata y asimismo en las larvas y ninfas de los insectos y los Collemboli y Acaridi adultos. 2) Si el animal es más complicado y voluminoso, la respiración simple tras el cutis no es suficiente por proveer los órganos internos. En esta segunda etapa del desarrollo el cambio de gases se produce también por la piel, pero el oxígeno está ligado a la hemoglobina y la sangre lo transporta a los órganos. Lo mismo vale para el ácido carbónico. Este estadio tienen los Vermes. (La consecuencia inmediata de este proceso respiratorio me parece ser la translocación del sistema nervioso ventral, porque el vaso sanguíneo debe necesariamente correr dorsal por el cambio de gases en un animal, que se arrastra por el suelo. Así el sistema nervioso es desalojado ventral en el abdomen, mientras permanece en la cabeza en su lugar primordial dorsal). 3) Cuando la piel llega a ser impenetrable por el aire, sea por el cuero, sea por la quitina, nuevos órganos han de ser creados para posibilitar el cambio de los gases. Esto se hace con la germinación de tubos, sea de la piel (tráqueas), sea de la mucosa del intestino (bronquios). Esto naturalmente no sirve sin un aparato de ventilación. 4) En los vertebrados es una bomba aspirante e impelente (Amphibia), que después es sustituida por una bomba solamente aspirante (Mamalia). 5) En los insectos existe solamente una bomba aspirante e impelente, que trae el oxígeno a los tejidos directamente, y totalmente distinto es el aparato para la eliminación del ácido carbónico. Aquí la comunicación primordial del sistema respiratorio y sanguíneo se ha mantenido, como en los Vertebrados. 6) La divergencia de estos dos grandes "Phyla" consiste principalmente, en que en los Vertebrados la respiración del cutis está reemplazada por la del intestino y que con esto han conservado la conexión total con la circulación sanguínea, mientras los insectos han conservado la respiración ectodermal con las tráqueas, pero han cortado en este respecto la conexión con la sangre, que no han retenido que por el transporte del ácido carbónico. Por esto han sido creados nuevos órganos de escape: los espiráculos. 7) Una forma primordial, de la cual se podría derivar Insectos y Vertebrados ha de ser más detrás de los Vermes. Debe ser un animal con una circulación sanguínea, que transporta los gases, pero que no ha desalojado el sistema nervioso. Resumen del desarrollo del sistema respiratorio en los Ateleceratas A. El arribo de oxígeno 1) La aspiración del aire en el cuerpo se efectúa: a) Por sacos dilatados y comprimidos por músculos (Scolopendra). b) Después la diferenciación del cuerpo en tórax y abdomen, con la reducción de extremidades, la respiración aprovecha de los segmentos abdominales para la inspiración. Los sacos respiratorios acaban de ser órganos de importancia primaria (Lepisma). c) Y se pierden en los demás insectos. Con la dilatación del abdomen se produce una presión menor, que aspira el aire. d) Una bomba singular en la coxa de la pulga. 2) El transporte del aire aspirado se efectúa después del cierre de los estigmas por una presión con los mismos órganos mencionados arriba. 3) El desarrollo de los estigmas es muy acentuado. a) Primordialmente un simple margen de quitina con un cojín interno es cerrado por músculos externos (Scolopendra, Cicadea) o bien indirectamente (Grillus). b) Una válvula automática según el principio de las botellas de aguas gaseosas, que cierran con una bola de vidrio. (Pulga). c) Finalmente el estigma es un aparato complicado con válvulas reguladas por músculos propios. (Hormigas, etc.). 4). La localización de los estigmas es: a)En todos los segmentos del cuerpo Scolopendra, b) Al menos de la cabeza (Pulex). c) Solamente en el abdomen (Los demás insectos). 5) Las tráqueas son netamente metaméricas y corren en paquetes a los órganos Scolopendra, Grillotalpa (Nussbaum), Campodea, Machilis (Grassi) Meloidi. b) Pulex posee también las tráqueas segmentales, que son pero unidas por anastomosis oblicuas intersegmentales. c) Finalmente existen troncos longitudinales, que están junto a los estigmas segmentales. (Acrididae, Hymenoptera, etc). 6) Cuando los troncos longitudinales aparecen, surgen una cantidad de órganos regulatorios. Se, refina el sistema traqueal con válvulas en forma de embudo, tubos delgados intercalados por un fluido constante del aire, músculos para la propulsión (corazones traqueales). Con esto las tráqueas primordialmente muy anchas pierden de calibre, porque con estos órganos regulatorios el aire es mejor dirigido. 7) Hay que distinguir entre los sacos respiratorios ventiladores de Scolopendra y Lepisma (en regreso) y los sacos intercalados en las tráqueas de los insectos volantes. Estos últimos son originalmente tráqueas ensanchadas con hilo espiral, que después es reemplazado por un estrato muscular. Estos sacos son depósitos de aire, que proveen la exigencia aumentada del oxígeno, durante el vuelo, y acumulan el aire rico de ázoe, que cambian después con aire normal por una verdadera expiración por los estigmas abiertos, regulados por válvulas con músculos. B. La evacuación del ácido carbónico 1) La conexión primordial entre la sangre y los gases de la respiración permanece inalterada entre la sangre y el ácido carbónico. 2) Con la quitinización del cutis han de ser construidos nuevos aparatos de escape, que he nombrado espiráculo. Parece que los estigmas toracales supérfluos han sido aprovechados y modificados. En los espiráculos no hay criba ni válvula. Falta la conexión con el sistema traqueal, pero siempre existe una comunicación con el vaso sanguíneo dorsal. 3) Primordialmente un saco abraza la aorta en cada segmento con una cámara de aire, que desemboca en ambos lados al exterior. (Scolopendra, Lepisma). 4) Después estos órganos de escape son limitados, en la pulga en la cabeza, las más veces a tres en los segmentos toracales (Mantis) y después, en los dos últimos segmentos toracales y el primer abdominal (Metanotum, Epinotum y Petiolum de las Hormigas). Si el animal es grueso, los espiráculos, confluyen en un impar (abeja, Langosta, Coleópteros, etc). 5) Por la protección del animal todos los espiráculos, como puntos débiles de la coraza están ocultos, sea por su sitio (abejas, Cicadas) o por camouflage (Libélulas, Mariposas, Myrmeleon) o son protegidas por láminas de quitina como el Scutellum u otros órganos de protección (Coleópteros, Chinches, Aethalión). 6) Comúnmente el ácido carbónico sale del cuerpo sin propulsión especial, solamente por la difusión del gas. 7) Como el ácido carbónico está ligado a la sangre, no es cierto. Diversas especies de células han sido descritas en la sangre, de las cuales no se conoce la función. Naturalmente podría ser el plasma el medio de transporte como en los Mamalia; cosa que a mi parecer es la más probable. Resumen 1) En la primera parte han sido descritos los órganos de escape por el ácido carbónico en conexión con la aorta, en representantes de cada orden de insectos. No hay insectos en que falten. 2) Sumergiendo un insecto con su tórax en agua de bario se comprobó que las burbujas de aire emitidas dorsal, son realmente ácido carbónico. El abdomen afuera del líquido permite la duración ilimitada de este experimento, porque los animales pueden inspirar libremente el oxígeno con el abdomen. 3) El oxígeno es empujado por el sistema traqueal directamente hasta los tejidos; y es consumido por las células. El ázoe superfluo del aire está acumulado en. los sacos respiratorios, que durante el vuelo suplen el oxígeno y después en el descanso el aire de los sacos, rico de ázoe y pobre de oxígeno es expirado por los estigmas, que poseen válvulas reguladas por músculos. 4) Después de una exposición del desarrollo filogenético de la respiración, se viene a la conclusión, que después que el cutis resulta impermeable por los gases (quitina o cuero), los Insectos conservan la respiración ectodermal con el sistema traqueal por el arribo de oxígeno, pero disuelven por este gas la conexión primordial con la sangre, que conservan por el transporte del ácido carbónico. Por esto se forman nuevos órganos de escape, los espiráculos. En los vertebrados, por el contrario, la conexión de la respiración con el sistema sanguíneo queda intacta en toda su extensión, pero la respiración cutánea es cambiada con la intestinal. Zusammenfassung 1) Im speciellen Teil wurde bei Vertretern jeder Insektenordnung dorsal gelegene Organe beschrieben, welche in Verbindung mit der Aorta stehen, und das Entweichen der Kohlensaeure aus dem Blut ermoeglichen. Sie fehlen nirgends. 2) Durch Eintauchen verschiedener Insekten in Barytwasser wurde nachgewiesen, dass die dorsal austretenden Luftblasen tatsaechlich Kohlensaeure sind. Die Tiere deren Hinterleib ausserhalb der Fluessigkeit blieb, die also ungehindert Sauerstoff einatnlen konnten, ueberlebten das Experiment ungeschaedigt. 3) Der Sauerstoff wird durch das Tracheensystem direkt den Geweben zugefuehrt und dort verbraucht. Der mitgefuehrte Stickstoff muss aus dern Koerper eliminiert werden, Waehrend des Fluges liefern die Luftsaecke Sauerstoff den Geweben und danach ist die Luft in diesen Reservesaecken Stickstoff-reich und Sauerstoff-arm. Sie wird dann durch rasche Abdominalbewegungen durch echtes Exspirium ausgeatmet durch die Stigmen, deren Klappen durch wilikuerliche Muskeln bedient werden. 4) Nach einem phylogenetischen Abriss der Entwicklung der Atemorgane, wird der Schluss gezogen, dass bei Undurchlaessigwerden des Integuments durch Verhornung oder Chitinisierung, die Insekten die Hautatmung im Tracheensystem beibehalten haben und dabei die urspruengliche Verbindueng mit dem Blutgefaess-system als Transportmittel fuer den Sauerstoff geloest, fuer die Koblensaeure aber beibehalten haben unter Neubildung von Abzugsorganen von der Aorta zur Aussenwelt. Die Wirbeltiere dagegen haben die Verbindung mit dem Blut als Transportmittel fuer die Atemgase voll beibehalten, dafuer die Hautatmung mit der Darmatmung vertauscht. |
|||||||||||||||||||||||||||||||||||||||||||||